ADVICE TO APPLICANTS
RESEARCH
HIGHLIGHTS
Lab director
Peter Chesson
Post-doctoral Associates
Lina Li
Graduate Students
Galen Holt
Chi Yuan
Yue (Max) Li
Pacifica Sommers
Simon Stump
Nick Kortessis
Recent PhDs
Megan Donahue
Anna Sears
Jessica Kuang
Adam Miller
Maureen Ryan
Recent Postdocs
Andrea Mathias
Jessica Kuang
Danielle Ignace
Undergraduate Students
Elieza Tang
Krista Cole
Friends of the Lab
Andrea Mathias
Adam Miller
Charlotte Lee
Brett Melbourne
Kendi Davies
Maureen Ryan
Scott Stark
Yun Kang
Graduation Photos
 |

Our study systems
Some of us are fond of studying communities of
annual plants. We find these in the deserts of Arizona and Eyre
Pennisula
(South Australia), in the vernal pools of the Central Valley of
California,
and in the coastal
swales near Bodega Bay. We also study
perennial herbs in the
understorey of Eucalypt forests in Australia. Some of us
are aquatic in orientation studying aquatic insects and amphibia in
ponds and streams, or invertebrates in
marine environments.
There are lots of systems that we think about
but do not collect data on ourselves, at least not right now. These
include coral
reef fish communities, tropical forests, and host-parasitoid systems.
Our tools
For studies of natural systems, we are
developing new classes of quantitative tools to test coexistence
mechanisms and hypotheses about dynamics of populations and communities
subject to spatial and temporal variation. Then we go out into
the field to count, measure, and manipulate.
For theoretical studies, we use math and
computers. We are fond
of probability and statistical theory. We also get into
differential equations
and Fourier transforms.
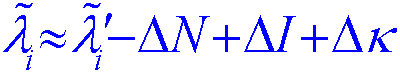
|
Our questions
Ecological
diversity
and its maintenance
The diversity of life is the wonder of the world. It inspires us, but
it challenges us to explain it too. Sadly also, diversity is under
threat on many fronts from human activities. To confront this threat we
need better understanding of the maintenance of diversity. The
diversity of life involves ecological diversity: different organisms do
different things in the
aid of their lives and their offspring. Some differences are subtle.
Some differences are gross. How do these differences arise, and what is
their role in maintaining diversity? How do the attributes of species
predict community change with environmental change?
General
testable community theory
To
understand diversity maintenance, we need theory for ecological
communities. It must be realistic enough to contain the key features
behind diversity maintenance in nature, it must be general enough to be
robust to variation in minor features, and it must be testable too.
Nature has many complexities, not the least of the which are the many
different species in any ecological community interacting with one
another in many different ways. The environments of organisms are
complex too. They vary greatly in space and time interacting with the
lives of organisms and their relationships with others. We try to rise
the challenge of this complex work, seeking theory that defines general
principles, and shows how to test them in specific contexts. We make it
quantitative, operational
and testable in practice. Then we go out and test it.
Variation and
scale
Ecological systems are characterized by
variability on all scales. Individual organisms vary one from another,
their environments vary every which way, populations fluctuate over
time and in space, communities vary and change in
composition. We ask how variation affects the way ecological systems
work. On the individual level, life histories may be adapted to
variation, ready to take advantage opportunities, but hedging against
uncertainties. Life-history properties, and the variation moulding
them,
have consequences for population dynamics and species interactions.
Population dynamics take different forms on different temporal and
spatial scales. Spatio-temporal niche differentiation arises from
taking advantage of opportunities in space and time. Species diversity
is promoted, interactions are stabilized, ecosystem functioning is
affected, and invasion resistance develops. We develop the mathematical
theory behind these ideas. We develop the methods to test them, and
then we go out and test them.

|

Our
Concepts

Research Highlights
Community
dynamics in time and space
Natural communities
inhabitat a world of great complexity in time and space. The physical
environment has many spatial complexities in its topography, hydrology
and soils. And it changes over time: organisms experience
varying weather and climatic changes on many scales interacting in a
variety of ways with spatial environmental varation. Various
processes, not just the physical environment, but also interactions
between organisms, cause their densities to vary greatly in space and
time too. It has often been hypothesized that these different
kinds of
variation have important roles in diversity maintenance. But what
are those roles? Development of mathematical models with such
complexities is not easy, and so it is perhaps not surprising that much
mathematical theory, even if it addresses density variation in time or
space, rarely includes variation in the physical environment. The
theory coming from this lab is different. We have embraced
environmental complexity, and gathered the mathematical tools necessary
to study it. The result is a comprehensive multiscaled theory of
diversity maintenance that includes the standard equilibrium theory as
a special case. This work has led to the recognition of several new
species coexistence mechanisms including the
storage effect, nonlinear
competitive variance, and fitness-density
covariance.
Multitrophic
diversity maintenance
The study of diversity maintenance mechanisms has
traditionally focused on competition, with predation mostly seen as
modifying what competition does. However, there is growing
evidence that predation and competition can have very similar effects
on the maintenance of species diversity. This similarity of action
relies on the observation by Bob Holt (University of Florida) that both
predation and competition can lead to mutually negative indirect
interactions between species. Of most importance, such mutually
negative indirect interactions apply not just between species but also
to different individuals within the same species. Theoretical
consequences of these facts mean that predation and competition each
have the potential to limit diversity or promote diversity. For
both predation and competition it all depends on the extent to which
within species interactions outway between species interactions.
When predation and competition have similar strengths for a given set
of
interacting species, the overall outcome for diversity |
maintenance may
represent a compromise between the separate tendencies of predation and
competition in the given circumstances. In other cases, new mechanisms
promoting diversity arise from the interaction between predation and
competiton. If one mechanism is much stronger than the other, the
tendency of the stronger mechanism prevails.
One major thrust within the lab explores these
various ideas in
general models and in models of specific systems, such as annual plant
communities. We combine them with other thrusts on the effects of
environmental variation in space and time, and we ask also how trophic
cascades affect the maintenance of diversity in any given trophic level.
Testing
species coexistence mechanisms
One of the biggest
challenges in community ecology is explaining
patterns of species diversity. We cannot hope to explain these patterns
unless we understand mechanisms of species coexistence. Although
there are numerous
hypotheses about species coexistence, few have been demonstrated in
nature with any degree of confidence.
Our theoretical
developments have recently led to powerful new ways of testing
species coexistence mechanisms. These tests use field
manipulations to test directly for mechanism functioning. Unlike
many other tests, which merely indicate consistency with mechanisms,
these tests are definitive, and thus provide high confidence that the
mechanism is working in the system. Most empirical studies in the
lab are using these new methods.
The
basis of these
new methods is the quantification of mechanism
strength and the functional components of mechanisms discussed
next.
Quantifying
mechanism strength in terms of
functional components
How do species coexistence
mechanisms work? And how do we know how strong they
are? Our lab has developed measures
of mechanism strength based on how much a mechanism
increases the rate of recovery of a species from low density. In
nature, species always fluctuate, and if they are to remain in a
community, they must recover from their low excursions. Focusing
on
these recovery rates, we have found ways of quantifying mechanism
strength in terms of the |
functional
components that define the
mechanism. This has led not only to a big increase in
understanding of
mechanism functioning, but also powerful
new
ways of testing mechanisms.
Multiple
mechanisms, mulitple scales
Most people expect that most natural communities
have multiple mechanisms of diversity maintenance in operation.
Moreover, these mechanisms are likely to work on a variety
of
scales. How do we understand systems with such complexities? The method
of
quantifying mechanism strength discussed above allows us to
partitioning
the recovery rate into the contributions from different mechanisms and
the contributions from
different scales in space
and time.
This allows
comparative contributions of the various applicable mechanisms to be
assessed,
facilitating understanding both in models and in nature.
The
scale transition
in theory and practice
Much progress has been made in recent years in the
development of models of communities subject to spatial and temporal
variability. So active is the work on the spatial front that it
is regarded as a new field, spatial ecology. But a daunting
challenge is how you test spatial ecology in the field. An
equally daunting challenge is how you achieve general understanding of
spatial and temporal mechanisms. Developments in this lab over
many years have led to a body of work called scale transition
theory,
which explains spatial and temporal ecology in terms of interactions
between nonlinear population processes and variation in space and
time. Of most importance, this work leads to ways of testing
spatial and temporal ecology, which are general, powerful and pratical.
Ecological
complexity
The work discussed here provides one route to understanding ecological
complexity. It takes a complex dynamic phenomenon and asks what
is it about this complexity that is relevant to critical questions like
species
coexistence, population
persistence, and population
stability. The results are statistical measures of complexity
that are functional predictors of the outcomes to these questions.
|
Peter Chesson
 Community
Processes
in Variable Environments
Community
Processes
in Variable Environments
The natural world is inherently variable. The
physical environment varies on all scales and so do the organisms
inhabiting it. Organisms
are adapted to this variability, not merely hedging against uncertainty
but
taking advantage of opportunities that variability brings. My main
interest is how the adaptation of organisms to variability promotes
species diversity and affects ecosystem functioning. I do this
primarily theoretically by developing
mathematical models using probability theory and the methods of
theoretical
statistics. Recent research topics have involved neighborhood
competition
models, community assembly, invasion resistance, multitrophic diversity
maintenance, life-history theory and the various
manifestations
of temporal and spatial niches, also known as the storage effect and
relative
nonlinearity of competiton. In addition, I am involved with field
projects
on annual plants and herbaceous perennial plants. Other systems that
are
foci of theorizing are coral reef communities, tropical forests, and
communities
in mediterranean climates.
Advice
to applicants...
Some recent publications
Chesson,
P. 2000. Mechanisms of maintenance of species diversity. Annual Review
of Ecology and Systematics 31, 343-66.
Chesson,
P. 2000. General theory of competitive coexistence in spatially varying
environments. Theoretical Population Biology 58, 211-237.
Chesson, P. 2001.
Metapopulations. Pp 161-176 in Encyclopedia of
Biodiversity, Vol 4, Simon A. Levin, ed, Academic Press.
Shea,
K., Chesson, P. 2002. Community ecology theory as a framework for
biological invasions. Trends in Ecology and Evolution 17, 170-176.
Chesson,
P.,
Peterson, A.G. 2002. The quantitative assessment of the benefits of
physiological integration in clonal plants. Evolutionary Ecology
Research 4, 1153–1176.
Chesson,
P. 2003. Quantifying and testing coexistence mechanisms arising from
recruitment fluctuations. Theoretical Population Biology 64,
345–357.
Chesson,
P., Gebauer, R. L. E., Schwinning, S., Huntly, N., Wiegand, K.,
Ernest, S. K. M., Sher, A., Novoplansky, A., and Weltzin,
J.F. 2004. Resource pulses, species interactions and diversity
maintenance in arid and semi-arid environments. Oecologia 141,
236 - 253.
Facelli,
J.M., Chesson, P., Barnes, N. 2005. Differences in seed biology of
annual plants in arid lands: a key ingredient of the storage
effect. Ecology 86, 2998-3006.
Chesson,
P., and Lee, C.T. 2005. Families of discrete kernels for modeling
dispersal. Theoretical Population Biology 67, 241-256.
Chesson,
P., Donahue, M., Melbourne, B., Sears, A. 2005. Scale
transition theory for understanding mechanisms in
metacommunities. In Holyoak, M, Leibold, M.A., Holt, R.D., eds,
Metacommunities: spatial dynamics and ecological communities, pp
279-306
Sears,
A.L.W., Chesson, P. 2007. New methods for quantifying the
spatial storage effect: an illustration with desert annuals.
Ecology 88, 2240-2247
Chesson,
P. 2008. Quantifying and testing species coexistence
mechanisms. Pp 119 – 164 in Valladares, F., Camacho, A., Elosegui, A.,
Estrada, M., Gracia, C., Senar, J.C., & Gili, J.M., ed. “Unity in
Diversity: Reflections on Ecology after the Legacy of Ramon Margalef.”
Chesson,
P, and Kuang, J.J. 2008. The interaction between predation and
competition. Nature 456, 235-238
All
publications
pchesson@u.arizona.edu
Back to top

Lina Li
Communities in varying environments
Coexistence in a temporally varying environment is
predicted to arise when species differ in their temporal patterns of
resource consumption. However, key to this outcome is coupling of
resource consumption and resource availability. High resource
consumption rates, due to favorable environmental conditions, are
expected to draw down resources, causing high competition. The
phenomenon is called covariance between environment and competition
(covEC). For coexistence to occur, covEC must be weaker for a species
perturbed to low density (an “invader”) compared to an unperturbed
species (a “resident”). However, these outcomes have not been
thoroughly investigated in models. Instead, the formulation of
models often implicitly includes them. Thus, the theory is
incomplete because there is no satisfactory theory of this key
component, covEC. Moreover, it has been suggested detecting covEC in
nature provides a powerful means of testing coexistence by the storage
effect. The phenomenon of covEC therefore requires a thorough
theoretical understanding. Here we extend the MacArthur
consumer-resource model to varying environments. The explicit
resource dynamics in the MacArthur model allow a theory of covEC to be
developed. Our approach was to develop quantitative measures of factors
contributing to covEC, and studying their effects in computer
simulations. We found that relative speeds of resource
consumption, consumer
dynamics and environmental variation have major effects on covEC.
Resource speed by far had the dominant effect and can be divided into
two major components, resource amplitude, and resource phase relative
to consumers. Resident covEC is maximal when resource amplitude
is greatest and phase relative consumers is zero, and this is achieved
when resources turn over rapidly, most strongly promoting coexistence
by the storage effect. These results allow us to address the
fundamental ideas of Hutchinson’s Paradox of the Plankton. While
they do show that seasonal environmental variation can promote
coexistence of consumers on limited resources, it produces very
different predictions about time scales of environmental change and
consumer dynamics compared with Hutchinson. Contrary to
Hutchinson’s predictions we find that slow consumer dynamics are
favorable to coexistence and do not lead to averaging of environment
fluctuations, nullifying their effects. Critical to coexistence is
factors strengthening covEC for residents, found here to be fast
resource dynamics, not intermediate consumer dynamics.
linali19800815@yahoo.com
Back to top
Galen Holt
 Empirical study of
ecological concepts
Empirical study of
ecological concepts
I am interested in population and community dynamics in
variable
environments, especially how environmental heterogeneity enhances
persistence. I have a field system in streams in the Catalina
Mountains, focusing on the persistence of various aquatic organisms in
pools. I have worked also on models and experiments for
understanding annual plant coexistence. Below I have included the
abstract of a talk I gave at the ESA 2008 on this topic.
Species-specific
germination speed and
coexistence in a desert annual plant community.
Desert annual pants are frequently the subjects of
coexistence studies with a focus on the role of environmental variation. Germination rates are observed to vary
substantially
over time forming the basis of the storage-effect coexistence mechanism. Laboratory germination studies have
reproduced some of this variation, but do not replicate the full range
of
conditions found in the field. Here we
discuss one aspect of field variation that potentially has a major
effect on
plant species coexistence, but has been neglected in the past.
Experimental and
observational data from the desert annual plants at our field site near
Portal,
AZ show distinct species-specific differences in the speed of
germination
following rainfall. This indicates an important and previously
unexplored
opportunity for covariance between competition and the environment, a
key
quantity for coexistence by the storage effect. Most seeds are at or
near the
surface, and so moisture at the soil surface is critical for
germination.
Duration of this moisture varies greatly due to duration, and spacing
of
rainfall events, and also due to solar radiation, temperature, humidity
and
wind following these events. Field observations and laboratory
experiments show
that the observed differential germination speed is strong enough to
lead to different
species dominating due to different durations of soil moisture. Thus,
variation
in duration of surface soil moisture potentially contributes
substantially
yearly variation in the species composition of the annual plant
community. Using
data on germination speed from field observations and germination
experiments
in the lab, we built a model linking fluctuations in surface soil
moisture and
species’ germination speeds to predict the crop of plants resulting
from
specific rainfall patterns. Our model implies that the interaction of
species-specific germination speed with variation in soil moisture
makes a
major contribution to the storage-effect coexistence mechanism in this
community.
For example, species that germinate quickly have greater opportunity to
take
advantage of short-duration rainfall and surface soil moisture, while
other
species require a longer period of surface soil moisture to
successfully
germinate, but may be superior competitors once germinated.
Incorporating the
interaction between germination speed and the duration and spacing of
rainfall
greatly expands the opportunities for species coexistence in this
community.
This increased understanding of the factors controlling community
composition
also provides a more refined understanding of the long-term stability
of
species composition.
gholt@email.arizona.edu
Back to top

Chi Yuan
Models of species coexistence in a
temporally varying environment
I study mathematical models to understand how environmental
variation affects species coexistence. So far, I have studied the
coexistence mechanism called
relatively nonlinear competitive variance
(aka relative nonlinearity)
in the lottery model. I found that the storage effect is a much
stronger mechanism than relative nonlinearity. However, relative
nonlinearity does have important effects if some species
have large adult death rates while other species have small adult
death rates. I want to study other models and other mechanisms in my
PhD work
 Yue (Max) Li
Yue (Max) Li
Diversity maintenance
mechanisms in different systems
I am interested in how the mechanisms of diversity
maintenance
differ between different systems, such as deserts and tropical
forests. Recently, I have been focusing on frequency-dependent
predation as a coexistence mechanism with a view to understanding how
it might be studied in different ecosystems.
liyue@u.arizona.edu
Back to top

Pacifica
Biodiversity Patterns
I am interested exploring
patterns
of biodiversity empirically and theoretically.
Currently, I am incorporating predator
avoidance
(indirect predator effects) into multi-trophic models of population
dynamics
to look at its effects on coexistence between similar species. I
am beginning empirical work with a community of aquatic insects in
mountain streams.
Back
to top

Simon
Effect
of pathogens on species
coexistence
I am fascinated by the question of what allows
intra-guild
competitors to coexist,
and hope to study this phenomenon both theoretically and empirically.
Right now the specific question I am asking is,"How do
parasites, disesases, and other
pathogens affect coexistence?" I want to understand when they promote
coexistence, and when (if ever) they undermine it. I am still
developing a research plan, but I think
my model organisms
will be desert annual plants and parasitic fungi.
 My
key interest is understanding
fluctuation-dependent coexistence mechanisms focusing on how the
mechanism, relatively nonlinear competitive variance workswith multiple
competitive factors.
My
key interest is understanding
fluctuation-dependent coexistence mechanisms focusing on how the
mechanism, relatively nonlinear competitive variance workswith multiple
competitive factors.
Friends of
the Chesson Lab
former members

Danielle Ignace
Physiological Ecology of Plant Invasions
I am studying the causes of the dramatic
irruption of the Eurasian invasive plant, Erodium cicutarium, in the San
Simon Valley of southeastern Arizona. It many areas, it is now
95% or more of
the winter annual plant biomass, up from a miniscule
presence away from roads in the 1980s. At the same time, native
winter annual plants have become very rare and are showing changes in
species composition. This new monoculture potentially has
profound implications for biodiversity, habitat quality, ecosystem
functioning and forage quality of the rangeland. My research applies a
physiological, ecosystem, community and theoretical approach to
understanding the characteristics associated with the irruption and
distribution of E. cicutarium
and its impacts for biodiversity in the San Simon Valley.
ddignace@email.arizona.edu
Back to top

Jessica
Kuang
Interactions
between Predation and Resource Competition
To
study problems in community ecology I
have been investigating mathematical models of species interactions
which take the form of nonlinear multidimensional Markov processes in
continuous space. The basic questions concern species coexistence,
which means finding conditions under which no species density converges
to zero with time. The particular mathematical techniques are invasion
analysis and quadratic approximations of the model. The
particular
ecological problem studied is the effect of a common enemy ("apparent
competition") on the coexistence of species that compete for
common resources. Biodiversity and coexistence are well studied by many
ecologists over the past century; however, this is the first study
which consider both resource and apparent competition in a stochastic
model.
Jessic
Kuang's website
jjkuang@u.arizona.edu
Back to top

Andrea Mathias
Non-equilibrium dynamics and species coexistence
Non-equilibrium dynamics of natural ecosystems
have
a major
role in generating and maintaining biodiversity. The main objective of
my
research is the mechanistic understanding of this role in both
ecological and evolutionary context. In particular I am interested in
life history evolution with frequency dependent selection in temporally
fluctuating and spatially heterogeneous environments.
So far I have studied the conditions for the
development of polymorphisms in initially monomorphic populations
leading to adaptive speciation, allowed by different regimes of spatial
and temporal heterogeneity (Mathias et al., 2001; Cheptou and Mathias,
2001; Mathias and Kisdi, 2002).
Currently I am studying the competitive
mechanisms
promoting coexistence in a multiple resource competition model of
phytoplankton species, in the context of a generalized theory (Chesson,
1994; Chesson and Huntly, 1997; Chesson, 2000a, b). In this model the
growth rate of the species is a non-linear function of the limiting
factors, which can result in endogenously created oscillations of
density. This mechanism has enormous stabilizing potential
for the coexistence of a surprisingly high number of species on few
resources.
My goal is to understand the details of the role of relative
nonlinearity
in species coexistence, which has not yet been investigated
sufficiently
in this case.
In the future I would like to extend the theory
of
species coexistence in competitive systems to life history evolution in
order to understand
the mechanisms that promote the coexistence of different life history
strategies
in temporally fluctuating and spatially heterogeneous environments.
Publications and
CV
amathias@u.arizona.edu
Back to top
Adam Miller
Spatial models of species coexistence
I study
mathematical models to understand how disturbance,
such
as
fire, affects species coexistence. Fire is known to be an
important factor in many ecological systems. In plant
communities, the combined effects of opening up space, burning biomass,
and releasing nutrients to the soil offer a variety of environmental
conditions. When burns are relatively frequent, and a stable burning
regime has persisted on evolutionary time scales, many species develop
specific responses to fires. Previous theory has shown that, when
species respond differently to environmental fluctuations, coexistence
can be promoted by certain types of interactions between environmental
and competitive factors. In this paper, we demonstrate how response
strategies to fire can form an axis of differentiation, allowing
species to stably coexist. In short, the action of differing fire
response strategies can generally provide a stabilizing storage
effect for the coexistence of species.
Maureen Ryan
Environmental
variation and species interactions
in
amphibian communities
I'm interested in how environmental variation promotes diversity within species and communities, and how mechanisms such as the storage effect that function through environmental variation can affect endangered species, in particular. My research focuses on a grassland salamander community in the Central Valley of California that includes the threatened California tiger salamander (Ambystoma californiense), hybrid tiger salamanders (A. californiense crossed with A. tigrinum mavortium, an introduced tiger salamander from the central US), and the California newt (Taricha torosa).
I'm using a combination of artificial pond experiments and field experiments to look at whether and how environmental variation promotes coexistence between the two native species, Ambystoma californiense and Taricha torosa, and how coexistence mechanisms operating in the native community are influenced by the addition of hybrid salamanders. Additionally, I'm looking at how environmental variation influences hybridization dynamics between A. cal iforniense and A. tigrinum mavortium and whether mechanisms associated with environmental variation can contribute to the maintenance of genetic polymorphism within hybridizing populations of tiger salamanders. Besides experiments, I'm using theoretical modeling and data from field surveys/sampling to investigate how dispersal and landscape structure interact with local community dynamics to influence species diversity and hybrid spread on a regional scale.
iforniense and A. tigrinum mavortium and whether mechanisms associated with environmental variation can contribute to the maintenance of genetic polymorphism within hybridizing populations of tiger salamanders. Besides experiments, I'm using theoretical modeling and data from field surveys/sampling to investigate how dispersal and landscape structure interact with local community dynamics to influence species diversity and hybrid spread on a regional scale.
From a conservation perspective, my work is motivated by an interest in how landscape alteration may influence amphibian diversity and species coexistence within extant communities, and in particular how common pond manipulations and the loss of vernal pools in the California Central Valley might influence California tiger salamander populations.

Anna Sears
Environmental variation and species interactions
Ecology is challenged to find simple ways to
understand the messiness of the natural world. I like to work from a
metacommunity
context, accepting that natural ecosystems contain a variety of
different
habitat types, and that this environmental variation affects the
interactions
of species within ecosystems. My PhD research specifically involves
using natural annual plant systems to test how variation in the
environment
affects species coexistence. I am also interested in how environmental
variation
affects food-web interactions, and generally interacts with other forms
of density
dependence. My field sites are in the Chihuahuan desert and at the
Bodega Marine
Reserve.
For my dissertation research, I studied how
spatial
variation in local density-dependent processes influences regional
dynamics. In collaboration with Marcel Holyoak, I investigated the
effects of local resource distribution, dispersal rate, and the
strength of density dependence on regional dynamics in a highly
manipulable microcosm system, in which we could track population
dynamics over time. Applying the same principles to a field system, I
studied Petrolisthes cinctipes,
a filter-feeding porcelain crab that lives in intertidal cobblefields
along the Pacific coast. Based out of Bodega Marine Laboratory, I
parameterized models of local density-dependence in this organism using
field and laboratory experiments. For Petrolisthes
cinctipes, the primary sources of density dependence are
gregarious settlement, competition, and predation. With Leah Akins, I
quantified the spatial variation that influences these process at two
scales: variation in P. cinctipes
density, predator abundance, and larval supply among rocks within sites
and among sites. By integrating information about spatial variation
into empirically-based models of local dynamics, we can predict what
local processes are most important when "scaling up" to regional
dynamics. For links to publications and Megan's CV, see her website.
 My
main
research
interest is population dynamics,
especially how small-scale heterogeneity affects dynamics at larger
scales. Understanding how spatial and temporal heterogeneity affect the
dynamics of populations and communities is fundamental to ecology.
Indeed, answers to the most prominent questions in ecology involve the
pervasive influence of heterogeneity. Why do species coexist? Why are
there so many species? Why do some species cycle whereas others don't?
How does habitat fragmentation affect extinction?
Why are some species more invasive than others? Spatial and temporal
variation in population densities and in the physical environment are
essential considerations in these questions. My current research and
research to date focuses on the effects of spatial heterogeneity.
Whereas empirical studies have typically focused on heterogeneity
exclusively, theory demonstrates that population dynamics are altered
by spatial heterogeneity only when heterogeneity interacts with
nonlinear demographic processes - the pillar for research in the
Chesson lab. The aim of my research is to develop empirical tools to
measure the interaction
of nonlinearity and heterogeneity and to use these tools to probe real
populations.
Thus, I tackle the important issue of how to integrate theory and
models
with data from real systems, to understand how the interaction of
spatial
heterogeneity and nonlinearity alters population dynamics. I use these
insights
to address important basic and applied questions in ecology.
My
main
research
interest is population dynamics,
especially how small-scale heterogeneity affects dynamics at larger
scales. Understanding how spatial and temporal heterogeneity affect the
dynamics of populations and communities is fundamental to ecology.
Indeed, answers to the most prominent questions in ecology involve the
pervasive influence of heterogeneity. Why do species coexist? Why are
there so many species? Why do some species cycle whereas others don't?
How does habitat fragmentation affect extinction?
Why are some species more invasive than others? Spatial and temporal
variation in population densities and in the physical environment are
essential considerations in these questions. My current research and
research to date focuses on the effects of spatial heterogeneity.
Whereas empirical studies have typically focused on heterogeneity
exclusively, theory demonstrates that population dynamics are altered
by spatial heterogeneity only when heterogeneity interacts with
nonlinear demographic processes - the pillar for research in the
Chesson lab. The aim of my research is to develop empirical tools to
measure the interaction
of nonlinearity and heterogeneity and to use these tools to probe real
populations.
Thus, I tackle the important issue of how to integrate theory and
models
with data from real systems, to understand how the interaction of
spatial
heterogeneity and nonlinearity alters population dynamics. I use these
insights
to address important basic and applied questions in ecology.
bamelbourne@ucdavis.edu
Kendi Davies
 The persistence
of species in patchy landscapes
The persistence
of species in patchy landscapes
My research focuses on testing and refining
theory
that makes predictions about the persistence of species in
heterogeneous landscapes, including metapopulation and related
spatio-temporal theory. So far, I have concentrated on the spatial and
temporal dynamics of populations and communities in landscapes that
have been modified by humans, through habitat fragmentation and
grazing. I am interested in how the life history characteristics of
species determine the response of species to landscape heterogeneity. I
use quantitative methods to test theory and have approached this work
using data from a large-scale, long-term field experiment for beetles
and data for a continental scale study of arid zone birds in Australia.
I am currently broadening this focus to explore
what
this same theory predicts about patterns of biological diversity in
naturally patchy
landscapes. I am focusing on the grassland plant community in
serpentine patches
in a non-serpentine matrix (in the University of California’s
McLaughlin Reserve).
Right now, I am particularly interested in spatial scale and the
relationship
between diversity and invasibility (Shea
and Chesson 2002).
Shea, K., Chesson, P. 2002. Community
ecology theory as a framework for biological invasions. Trends in
Ecology and Evolution 17, 170-176.
Some recent publications
Davies, K.F., Chesson, P., Harrison, S., Inouye, B.D.,
Melbourne,
B.A., Rice, K.J. 2005. Spatial heterogeneity explains the scale
dependence of the native-exotic diversity relationship. Ecology
86, 1602-1610.
Davies, K. F., C. R. Margules, and J. F.
Lawrence.
2004. A synergistic effect puts rare,
specialized species at greater risk of extinction. Ecology 85: 265-271.
Davies, K. F., B. A. Melbourne, C. R.
Margules,
and
J. F. Lawrence (in press).
Metacommunity structure influences the stability of local beetle
communities. In M.
Holyoak, M. A. Leibold and R. D. Holt eds. Metacommunities: Spatial
Dynamics and
Ecological Communities. Chicago, University of Chicago Press.
Davies, K. F., B. A. Melbourne, and C. R.
Margules.
2001. Effects
of within- and between-patch processes on community dynamics in a
fragmentation experiment. Ecology 82: 1830-1846.
Davies, K. F., C. Gascon, and C. R. Margules.
2001.
Habitat Fragmentation: consequences, management and future research
priorities. Pages 81-97 in M. E. Soulé and G. H. Orians,
editors. Conservation Biology: Research Priorities for the Next Decade.
Island Press, Washington.
Davies, K. F., C. R. Margules, and J. F.
Lawrence.
2000. Which
traits of species predict population declines in experimental forest
fragments? Ecology 81: 1450-1461.
Davies, K. F., and C. R. Margules. 1998.
Effects
of
habitat fragmentation on carabid beetles: experimental evidence.
Journal of Animal Ecology 67: 460-471.
Charlotte
Lee
Theory of multispecies
interactions; human-environment interactions
I study the theory of multispecies interactions,
with specific focus on
the matrix of coefficients for all pairwise interactions in a
community.
Specific projects include: 1) Determining relationships between
different
patterns of direct and indirect competitive (or facilitative)
interactions
and the structure of model community matrices; 2) determining patterns
of
shared resource use that result in specific community matrices; and 3)
developing techniques to do 1) and 2). I also study human-environment
interactions using theory from ecosystem ecology and demography.
Charlotte.Lee@stanford.edu
Friends of the Chesson Lab
collaborators from other labs

Scott Stark
Diversity maintenance in tropical plant communities
The classic gap hypothesis suggests
that canopy gaps promote forest
diversity by favoring the regeneration of tree species with high light
requirements (i.e., shade intolerant species). However, whether gaps
act to maintain forest diversity remains an unanswered question. We
argue that canopy gaps can promote coexistence between tree species
through the ‘storage effect.’ Certain combinations of environmental
conditions and species’ traits can advantage species when they are rare
and promote their long-term persistence in the community through the
‘storage effect.’ These combinations are likely to be found in canopy
gaps. For example, gaps (and subsequently light environments) vary in
both space and time and tree species appear to be differentiated in
terms of their ability to tolerate shade and respond to increases in
light. Furthermore, when gap environments are favorable to abundant
species their ability to increase in dominance is likely limited
(relative to rare species) because of increased within-species
competition (i.e., covariance of environmental favorability and
competition). Finally, rare species will likely persist even when
favorable gap conditions are infrequent because adult trees are
long-lived. We evaluate the ‘gap storage hypothesis’ with quantitative
analyses that incorporate realistic patterns of variation in light
environments (i.e., gaps) and trade-offs between species ability to
tolerate shade and grow in high light. We show that species
differentiated along this trade-off may coexist via the ‘storage
effect.’ We conclude that the ‘storage effect’ may act to maintain
forest diversity because of canopy gaps.

Yun Kang
In collaborating with the Chesson Lab,
I am particularly interested in
relative nonlinearity as a coexistence
mechanism. I work in other areas of mathematical biology modeling,
including nonlinear ecological models, stoichiometric ecology,
spatial ecology, and epidemiology. I have also studied random,
inhomogeneous, and dynamic graphs.
Yun.Kang@asu.edu







